The Fact That Cones Are Most Sensitive to Either the Colour Red Green or Blue Supports
A special property of the cone system is color vision. Perceiving color allows humans (and many other animals) to discriminate objects on the basis of the distribution of the wavelengths of light that they reflect to the eye. While differences in luminance are often sufficient to distinguish objects, color adds another perceptual dimension that is especially useful when differences in luminance are subtle or nonexistent. Color obviously gives us a quite different way of perceiving and describing the world we live in.
Unlike rods, which contain a single photopigment, there are three types of cones that differ in the photopigment they contain. Each of these photopigments has a different sensitivity to light of different wavelengths, and for this reason are referred to as "blue," "green," and "red," or, more appropriately, short (S), medium (M), and long (L) wavelength cones, terms that more or less describe their spectral sensitivities (Figure 11.12). This nomenclature implies that individual cones provide color information for the wavelength of light that excites them best. In fact, individual cones, like rods, are entirely color blind in that their response is simply a reflection of the number of photons they capture, regardless of the wavelength of the photon (or, more properly, its vibrational energy). It is impossible, therefore, to determine whether the change in the membrane potential of a particular cone has arisen from exposure to many photons at wavelengths to which the receptor is relatively insensitive, or fewer photons at wavelengths to which it is most sensitive. This ambiguity can only be resolved by comparing the activity in different classes of cones. Based on the responses of individual ganglion cells, and cells at higher levels in the visual pathway (see Chapter 12), comparisons of this type are clearly involved in how the visual system extracts color information from spectral stimuli. Despite these insights, understanding of the neural mechanisms that underlie color perception has been elusive (Box D).

Figure 11.12
Color vision. The absorption spectra of the four photopigments in the normal human retina. The solid curves indicate the three kinds of cone opsins; the dashed curve shows rod rhodopsin for comparison. Absorbance is defined as the log value of the intensity (more...)
![]()
Box D
The Importance of Context in Color Perception.
Much additional information about color vision has come from studies of individuals with abnormal color detecting abilities. Color vision deficiencies result either from the inherited failure to make one or more of the cone pigments or from an alteration in the absorption spectra of cone pigments (or, rarely, from lesions in the central stations that process color information; see Chapter 12). Under normal conditions, most people can match any color in a test stimulus by adjusting the intensity of three superimposed light sources generating long, medium, and short wavelengths. The fact that only three such sources are needed to match (nearly) all the perceived colors is strong confirmation of the fact that color sensation is based on the relative levels of activity in three sets of cones with different absorption spectra. That color vision is trichromatic was first recognized by Thomas Young at the beginning of the nineteenth century (thus, people with normal color vision are called trichromats). For about 5–6% of the male population in the United States and a much smaller percentage of the female population, however, color vision is more limited. Only two colors of light are needed to match all the colors that these individuals can perceive; the third color category is simply not seen. Such dichromacy, or "color blindness" as it is commonly called, is inherited as a recessive, sex-linked characteristic and exists in two forms: protanopia, in which all color matches can be achieved by using only green and blue light, and deuteranopia, in which all matches can be achieved by using only blue and red light. In another major class of color deficiencies, all three light sources (i.e., short, medium, and long wavelengths) are needed to make all possible color matches, but the matches are made using values that are significantly different from those used by most individuals. Some of these anomalous trichromats require more red than normal to match other colors (protanomalous trichromats); others require more green than normal (deuteranomalous trichromats).
Jeremy Nathans and his colleagues at Johns Hopkins University have provided a deeper understanding of these color vision deficiencies by identifying and sequencing the genes that encode the three human cone pigments (Figure 11.13). The genes that encode the red and green pigments show a high degree of sequence homology and lie adjacent to each other on the X chromosome, thus explaining the prevalence of color blindness in males. In contrast, the blue-sensitive pigment gene is found on chromosome 7 and is quite different in its amino acid sequence. These facts suggest that the red and green pigment genes evolved relatively recently, perhaps as a result of the duplication of a single ancestral gene; they also explain why most color vision abnormalities involve the red and green cone pigments.
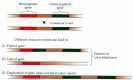
Figure 11.13
Many deficiencies of color vision are the result of genetic alterations in the red or green cone pigments due to the crossing over of chromosomes during meiosis. This recombination can lead to the loss of a gene, the duplication of a gene, or the formation (more...)
Human dichromats lack one of the three cone pigments, either because the corresponding gene is missing or because it exists as a hybrid of the red and green pigment genes (see Figure 11.13). For example, some dichromats lack the green pigment gene altogether, while others have a hybrid gene that is thought to produce a red-like pigment in the "green" cones. Anomalous trichromats also possess hybrid genes, but these elaborate pigments whose spectral properties lie between those of the normal red and green pigments. Thus, although most anomalous trichromats have two distinct sets of long-wavelength cones (one normal, one hybrid), there is more overlap in their absorption spectra than in normal trichromats, and thus less difference in how the two sets of cones respond to a given wavelength (with resulting anomalies in color perception).
Source: https://www.ncbi.nlm.nih.gov/books/NBK11059/
0 Response to "The Fact That Cones Are Most Sensitive to Either the Colour Red Green or Blue Supports"
Post a Comment